Biodiversità e diversità criptica in ambiente marino. Il codice a barre degli organismi viventi
Dott. Gabriele Procaccini,
Ricercatore della Stazione Zoologica A. Dohrn

Il mare ospita una straordinaria quantità di forme viventi.
La sua apparente uniformità contrasta con la reale diversificazione degli ambienti, che è ciò che ha permesso che organismi appartenenti a tutti i maggiori raggruppamenti del mondo vivente possano occupare nicchie diverse, svolgere funzioni ecologiche diversificate, interagire a livelli diversi ed in modo diverso con l’ambiente fisico, chimico e biologico che li circonda.
La ricorrenza del termine “diverso” non è casuale. Quello che è fondamentale sottolineare a questo punto è infatti la estrema diversità esistente in mare, sia a livello macroscopico, e quindi facilmente osservabile ad occhio nudo, che microscopico, all’interno di una goccia d’acqua o di un vetrino di microscopio. Questa enorme varietà di forme e di funzioni è stata classificata all’interno di schemi definiti; attraverso criteri che si basano essenzialmente sulle affinità di forme e sulla continuità riproduttiva è stato così creato il concetto di specie.
Qualunque persona che osservi l’ambiente marino attraverso i vetri di un acquario o quelli di una maschera è in grado di riconoscere un certo numero di specie di pesci, di invertebrati o di alghe e di assegnargli dei nomi, da quelli comuni a quelli scientifici.
Il nome scientifico è costituito da due parole e racchiude in se una doppia informazione, la identificazione della specie e la appartenenza al gruppo tassonomico di rango più alto, il genere. Il ‘sarago pizzuto’ sarà così Diplodus puntazzo, definendone la sua identità specifica e la sua appartenenza al genere Diplodus, cosa che indica l’esistenza di forti affinità con le altre specie appartenenti allo stesso genere.
Questo esercizio di classificazione del mondo vivente e questo tentativo di inscrivere tutte le forme in categorie di rango crescente in base alle affinità morfologiche ed alle storie evolutive, è iniziato agli albori della osservazione del mondo naturale da parte dell’uomo ma ha ricevuto la sua formalizzazione nel 1735 da parte dello studioso svedese Carolus Linnaeus che nel suo trattato ‘Systema Natura’ usò per la prima volto il doppio nome per definre una singola specie. Da quel momento in poi tutti gli organismi sono stati classificati in specie, le specie raggruppate in Generi, i Generi in Famiglie e così via fino ai livelli filetici più alti che rappresentano i Phyla e poi i Regni.
A questo punto, perchè il discorso seguente sia di facile comprensione, è importante introdurre un concetto nuovo, quello di affinità evolutiva. Utilizzando di nuovo l’esempio del sarago potremo dire che la specie D. puntazzo e le altre specie appartenenti allo stesso genere possono essere raggruppate grossolanamente in base alle loro caratteristiche morfologiche ma perchè siano realmente ascrivibili allo stesso genere devono condividere un antenato comune, dal quale esse si sono originate. E questo antenato deve essere più vicino, dal punto di vista evolutivo, a tutte le specie del genere di quanto lo sia a specie di generi diversi.
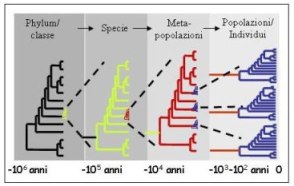
I raggruppamenti tassonomici vengono identificati attraverso l’origine degli organismi da antenati comuni
Fino a questo punto sembra tutto semplice ed estremamente logico. In realtà lo studio della biodiversità (la diversità delle componenti biologiche dei sistemi naturali) racchiude una serie di problemi e di difficoltà metodologiche. Come ricercare il famoso antenato comune? Come distinguere fra caratteri morfologici che sono simili per similarità adattative da quelli che derivano da comuni storie evolutive? Come distinguere fra specie diverse che sono morfologicamente simili se non identiche? Come tracciare il reale confie esistente fra specie diverse in quei casi in cui la specie stessa presenti un forte polimorfismo morfologico ed un gradiente di variabilità dei caratteri? Come valutare il livello di diversità esistente all’interno di una singola specie e la sua importanza rispetto alla diversità esistente fra specie distinte? Come stabilre la storia evolutiva delle popolazioni, la loro espansione e la loro capacità di persistenza?
La difficoltà a fornire una chiara risposta a molte di queste domande ha rappresentato il limite reale dell’analisi tassonomica classica, basata solo sullo studio della morfologia degli organismi. La rivoluzione più significativa in questo campo è rappresentata dall’avvento delle tecniche di analisi diretta delle proteine e degli acidi nucleici, ed in particolare del DNA.
Si tratta di una rivoluzione abbastanza recente ed infatti i primi studi che affrontavano problematiche ecologiche e tassonomiche attraverso l’analisi dei polimorfismi proteici, sono dei primi anni ’60. Successivamente, dagli inizi degli anni ’70 fino ad oggi, gli enormi e rapidi progressi della biologia molecolare hanno messo a disposizione dell’ecologo, del tassonomo e dell’evoluzionista una serie di strumenti sempre più sofisticati per guardare direttamente alla conformazione ed alla composizione della doppia elica del DNA, e per valutarne le affinità e le differenze fra gli organismi viventi.
Questi tipi di approcci si basano sull’assunzione che organismi filogeneticamente vicini (i nostri saraghi del genere Diplodus dell’esempio precedente) avranno fra di loro maggiori affinità nella successione di basi azotate che compongono il loro DNA rispetto a quelle esistenti fra specie appartenenti a generi diversi. E’ facile intuire che questo tipo di approfondimento ha dato la possibilità di guardare alla biodiversità da un punto di vista completamente diverso e con un livello di approfondimento impensabile fino a poco più della metà del secolo scorso. L’osservazione del mondo naturale e la sua classificazione in categorie e comparti si è svincolata infatti dalla sola analisi del morfotipo (l’aspetto morfologico dell’organismo) per passare alla analisi del genotipo (la caratterizzazione genetica dell’organismo) attraverso la caratterizzazione di quelle zone del DNA più svincolate dalle pressioni selettive dell’ambiente esterno. Ogni individuo, ogni specie, ogni genere e via via andando sempre più su nella scale dei livelli tassonomici, saranno caratterizzati da un ‘DNA fingerprinting’ specifico, da un codice a barre che permetterà di distinguerli dagli altri e di stabilirne relazioni evolutive, affinità filogenetiche ed appartenenze agli stessi gruppi tassonomici.

Ogni specie è caratterizzata da una sequenza diversa di regioni specifiche del DNA
Lo studio della filogenesi e della tassonomia molecolare degli organismi marini ha permesso di scoprire che i livelli di diversità precedentemente accreditati in alcuni casi sottostimavano ed in altri travisavano la reale diversità esistente.
La caratterizzazione molecolare di specie di difficile analisi morfologica, come gli esemplari microscopici del fito e dello zooplankton, sta portando alla scoperta di una serie di ‘specie criptiche’, morfologicamente indistinguibili ma filogeneticamente distanti, prima del tutto ignorate.
Lo studio di alcune specie di macroalghe ha confermato in alcuni casi che specie diverse in realtà non sono altro che fasi diverse del ciclo vitale della stessa specie.
Lo studio, infine, della diversità genetica presente all’interno della stessa specie costituisce il campo della filogeografia e della genetica di popolazione.
Attraverso questi studi, del nostro sarago potremo scoprire anche i rapporti “di parentela” esistenti fra individui appartenenti alla stessa popolazione o a popolazioni distinte, la sua capacità di dispersione e di spostamento fra località diverse, la eventuale capacità di reazione delle sue popolazioni alla presenza di uno stress esterno ed infine aiutare nella definizione delle corrette strategie di gestione e di salvaguardia.
foto di Archivio SZN – Laboratorio di Ecologia del Benthos – si ringrazia per la gentile concessione.
Lascia un Commento
Ischia, l'invasione dell'alga rossa su BIR
 Un contributo all'ampliamento della Conoscenza Ecologica Locale.
Siamo lieti di [.....]
Un contributo all'ampliamento della Conoscenza Ecologica Locale.
Siamo lieti di [.....]
 il video del salvataggio effettuato il 20/06 ad Ischia [.....]
il video del salvataggio effettuato il 20/06 ad Ischia [.....]
L'invasione negletta: L. lallemandii ad Ischia
 Lophocladia lallemandii è un'alga invasiva. Segnalazione dell'invasione nel sito di [.....]
Lophocladia lallemandii è un'alga invasiva. Segnalazione dell'invasione nel sito di [.....]
le fosse della neve a ischia - il video
 Scenari evocativi di un passato non molto lontano, quelli immortalati [.....]
Scenari evocativi di un passato non molto lontano, quelli immortalati [.....]
ottobre nel mare di Ischia - il video
 in una giornata di inizio ottobre siamo usciti in mare [.....]
in una giornata di inizio ottobre siamo usciti in mare [.....]
Pesci pappagallo ad Ischia: le foto
 le foto scattate dall'Associazione Nemo a quattro pesci pappagallo nell'estate [.....]
le foto scattate dall'Associazione Nemo a quattro pesci pappagallo nell'estate [.....]
Conoscenza Ecologica Locale e Cittadinanza Scientifica
 [articolo rivisto e ampliato sul notiziario SIBM 67/2015]
Ci fu un tempo [.....]
[articolo rivisto e ampliato sul notiziario SIBM 67/2015]
Ci fu un tempo [.....]
 La stagione turistica è terminata. Partiti gli ultimi autobus turistici [.....]
La stagione turistica è terminata. Partiti gli ultimi autobus turistici [.....]
Sereno Variabile con Nemo: il video
 La puntata della trasmissione di RAI 2 "Sereno Variabile", andata [.....]
La puntata della trasmissione di RAI 2 "Sereno Variabile", andata [.....]
Barchette arenate a Sant'Angelo: le foto.
 Le Barchette di San Pietro: un blu che affascina. Scopri [.....]
Le Barchette di San Pietro: un blu che affascina. Scopri [.....]
EcoGenesi: una goccia nel mare provoca seri danni
 - mappa dei punti raccolta oli esausti -
Avete mai pensato [.....]
- mappa dei punti raccolta oli esausti -
Avete mai pensato [.....]
CAMERA IPERBARICA CRISTIANO IELASI
 QUEI 500MILA EURO “SCOMPARSI” - LI AVEVA PROMESSI E POI [.....]
QUEI 500MILA EURO “SCOMPARSI” - LI AVEVA PROMESSI E POI [.....]
 Cronaca di un incontro pubblico di Luca Tiberti.
Il giorno 25 [.....]
Cronaca di un incontro pubblico di Luca Tiberti.
Il giorno 25 [.....]
| |
|
|

![]()



 |
 |
 |
 |
©2009-2022 - Associazione Nemo per la Diffusione della Cultura del Mare. Tutti i diritti riservati. Foto (se non specificato): Iacono B., Iacono G., Mattera F., Tiberti L.
| Sede Operativa estiva: borgo di Sant'Angelo, via Sant'Angelo 85 - 80070, Serrara Fontana (Isola d'Ischia, Napoli, Italia) guarda la mappa su google maps | ||||||||
| telefono: +393661270197 | ||||||||
| mail: info@nemoischia.it | ||||||||
| Sede Legale: via Prov.le Panza Succhivo, 26 - 80070, Serrara Fontana (Isola d'Ischia, Napoli, Italia) | ||||||||
| Codice Fiscale e P.IVA: 05089171218 | ||||||||
| IBAN: IT93S 03069 09606 100000018878 | ||||||||
| Posta Elettronica Certificata (PEC): nemoischia@pec.it | ||||||||
Nemo è partner di:
|
translate this page: | |||||||




|
|
|
